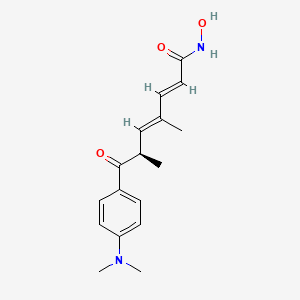
trichostatin A
Trichostatin is a lipid of Polyketides (PK) class. Trichostatin is associated with abnormalities such as Dentatorubral-Pallidoluysian Atrophy, PARAGANGLIOMAS 3, abnormal fragmented structure, Disintegration (morphologic abnormality) and Hyperostosis, Diffuse Idiopathic Skeletal. The involved functions are known as Acetylation, Cell Differentiation process, histone modification, Gene Silencing and Transcriptional Activation. Trichostatin often locates in CD41a, Hematopoietic System, Chromatin Structure, Blood and Endothelium. The associated genes with Trichostatin are SPI1 gene, CELL Gene, Chromatin, CXCR4 gene and DNMT1 gene. The related lipids are Butyrates, Promega, butyrate, Lipopolysaccharides and Steroids. The related experimental models are Knock-out, Mouse Model, Xenograft Model and Cancer Model.
Cross Reference
Introduction
To understand associated biological information of trichostatin A, we collected biological information of abnormalities, associated pathways, cellular/molecular locations, biological functions, related genes/proteins, lipids and common seen animal/experimental models with organized paragraphs from literatures.
What diseases are associated with trichostatin A?
trichostatin A is suspected in Infection, Morphologically altered structure, Ureteral obstruction, Photosensitization, Atherosclerosis, Hypertrophic Cardiomyopathy and other diseases in descending order of the highest number of associated sentences.
Related references are mostly published in these journals:
| Disease | Cross reference | Weighted score | Related literature |
|---|
Possible diseases from mapped MeSH terms on references
We collected disease MeSH terms mapped to the references associated with trichostatin A
PubChem Associated disorders and diseases
What pathways are associated with trichostatin A
Lipid pathways are not clear in current pathway databases. We organized associated pathways with trichostatin A through full-text articles, including metabolic pathways or pathways of biological mechanisms.
Related references are published most in these journals:
| Pathway name | Related literatures |
|---|
PubChem Biomolecular Interactions and Pathways
Link to PubChem Biomolecular Interactions and PathwaysWhat cellular locations are associated with trichostatin A?
Visualization in cellular structure
Associated locations are in red color. Not associated locations are in black.
Related references are published most in these journals:
| Location | Cross reference | Weighted score | Related literatures |
|---|
What functions are associated with trichostatin A?
Related references are published most in these journals:
| Function | Cross reference | Weighted score | Related literatures |
|---|
What lipids are associated with trichostatin A?
Related references are published most in these journals:
| Lipid concept | Cross reference | Weighted score | Related literatures |
|---|
What genes are associated with trichostatin A?
Related references are published most in these journals:
| Gene | Cross reference | Weighted score | Related literatures |
|---|
What common seen animal models are associated with trichostatin A?
Mouse Model
Mouse Model are used in the study 'Regulation of minichromosome maintenance gene family by microRNA-1296 and genistein in prostate cancer.' (Majid S et al., 2010), Mouse Model are used in the study 'Reversal of hypermethylation and reactivation of p16INK4a, RARbeta, and MGMT genes by genistein and other isoflavones from soy.' (Fang MZ et al., 2005) and Mouse Model are used in the study 'Histone deacetylase 3 mediates allergic skin inflammation by regulating expression of MCP1 protein.' (Kim Y et al., 2012).
Xenograft Model
Xenograft Model are used in the study 'Histone deacetylase inhibitors induce growth arrest and differentiation in uveal melanoma.' (Landreville S et al., 2012), Xenograft Model are used in the study 'Extended treatment with physiologic concentrations of dietary phytochemicals results in altered gene expression, reduced growth, and apoptosis of cancer cells.' (Moiseeva EP et al., 2007) and Xenograft Model are used in the study 'Retinoic acid and the histone deacetylase inhibitor trichostatin a inhibit the proliferation of human renal cell carcinoma in a xenograft tumor model.' (Touma SE et al., 2005).
Cancer Model
Cancer Model are used in the study 'Plasma pharmacokinetics and metabolism of the histone deacetylase inhibitor trichostatin a after intraperitoneal administration to mice.' (Sanderson L et al., 2004).
Related references are published most in these journals:
| Model | Cross reference | Weighted score | Related literatures |
|---|
NCBI Entrez Crosslinks
All references with trichostatin A
Download all related citations| Authors | Title | Published | Journal | PubMed Link |
|---|---|---|---|---|
| Mishra DR et al. | Identification of Critical Elements for Regulation of Inorganic Pyrophosphatase (PPA1) in MCF7 Breast Cancer Cells. | 2015 | PLoS ONE | pmid:25923237 |
| Yang X et al. | CDKN1C (p57) is a direct target of EZH2 and suppressed by multiple epigenetic mechanisms in breast cancer cells. | 2009 | PLoS ONE | pmid:19340297 |
| Liu W et al. | HDAC6 regulates epidermal growth factor receptor (EGFR) endocytic trafficking and degradation in renal epithelial cells. | 2012 | PLoS ONE | pmid:23152903 |
| Li L et al. | In Vivo Screening Using Transgenic Zebrafish Embryos Reveals New Effects of HDAC Inhibitors Trichostatin A and Valproic Acid on Organogenesis. | 2016 | PLoS ONE | pmid:26900852 |
| Bennett KL et al. | AP-2alpha induces epigenetic silencing of tumor suppressive genes and microsatellite instability in head and neck squamous cell carcinoma. | 2009 | PLoS ONE | pmid:19742317 |
| Tseng AS et al. | HDAC activity is required during Xenopus tail regeneration. | 2011 | PLoS ONE | pmid:22022609 |
| Zhao TC et al. | HDAC inhibition elicits myocardial protective effect through modulation of MKK3/Akt-1. | 2013 | PLoS ONE | pmid:23762381 |
| de Souza CF et al. | Mining gene expression signature for the detection of pre-malignant melanocytes and early melanomas with risk for metastasis. | 2012 | PLoS ONE | pmid:22984562 |
| Otsuji TG et al. | Dynamic link between histone H3 acetylation and an increase in the functional characteristics of human ESC/iPSC-derived cardiomyocytes. | 2012 | PLoS ONE | pmid:22984602 |
| Liu H et al. | The Smn-independent beneficial effects of trichostatin A on an intermediate mouse model of spinal muscular atrophy. | 2014 | PLoS ONE | pmid:24984019 |
| Sar P et al. | 3, 3'5 Triiodo L thyronine induces apoptosis in human breast cancer MCF-7 cells, repressing SMP30 expression through negative thyroid response elements. | 2011 | PLoS ONE | pmid:21687737 |
| Kim HR et al. | Aberrant Expression of TIMP-2 and PBEF Genes in the Placentae of Cloned Mice Due to Epigenetic Reprogramming Error. | 2016 | PLoS ONE | pmid:27855185 |
| Zou JY and Crews FT | Release of neuronal HMGB1 by ethanol through decreased HDAC activity activates brain neuroimmune signaling. | 2014 | PLoS ONE | pmid:24551070 |
| Gou Z et al. | Epigenetic modification of TLRs in leukocytes is associated with increased susceptibility to Salmonella enteritidis in chickens. | 2012 | PLoS ONE | pmid:22438967 |
| Tian Y et al. | Ubiquitin B in cervical cancer: critical for the maintenance of cancer stem-like cell characters. | 2013 | PLoS ONE | pmid:24367661 |
| Amoêdo ND et al. | Energy metabolism in H460 lung cancer cells: effects of histone deacetylase inhibitors. | 2011 | PLoS ONE | pmid:21789245 |
| Jagirdar K et al. | The NR4A2 nuclear receptor is recruited to novel nuclear foci in response to UV irradiation and participates in nucleotide excision repair. | 2013 | PLoS ONE | pmid:24223135 |
| Liu Y et al. | Expression loss and revivification of RhoB gene in ovary carcinoma carcinogenesis and development. | 2013 | PLoS ONE | pmid:24223801 |
| Reeves ME et al. | Evidence that RASSF1C stimulation of lung cancer cell proliferation depends on IGFBP-5 and PIWIL1 expression levels. | 2014 | PLoS ONE | pmid:25007054 |
| Anantharaju PG et al. | Induction of colon and cervical cancer cell death by cinnamic acid derivatives is mediated through the inhibition of Histone Deacetylases (HDAC). | 2017 | PLoS ONE | pmid:29190639 |
| Gurgul A et al. | Evaluation of changes arising in the pig mesenchymal stromal cells transcriptome following cryopreservation and Trichostatin A treatment. | 2018 | PLoS ONE | pmid:29390033 |
| Spin JM et al. | Chromatin remodeling pathways in smooth muscle cell differentiation, and evidence for an integral role for p300. | 2010 | PLoS ONE | pmid:21179216 |
| Estella C et al. | Inhibition of histone deacetylase activity in human endometrial stromal cells promotes extracellular matrix remodelling and limits embryo invasion. | 2012 | PLoS ONE | pmid:22291969 |
| Palazzese L et al. | Nuclear quiescence and histone hyper-acetylation jointly improve protamine-mediated nuclear remodeling in sheep fibroblasts. | 2018 | PLoS ONE | pmid:29543876 |
| Shakèd M et al. | Histone deacetylases control neurogenesis in embryonic brain by inhibition of BMP2/4 signaling. | 2008 | PLoS ONE | pmid:18628975 |
| Halaban R et al. | Integrative analysis of epigenetic modulation in melanoma cell response to decitabine: clinical implications. | 2009 | PLoS ONE | pmid:19234609 |
| Terhune SS et al. | Human cytomegalovirus UL29/28 protein interacts with components of the NuRD complex which promote accumulation of immediate-early RNA. | 2010 | PLoS Pathog. | pmid:20585571 |
| Dar RD et al. | Transcriptional burst frequency and burst size are equally modulated across the human genome. | 2012 | Proc. Natl. Acad. Sci. U.S.A. | pmid:23064634 |
| Ohno Y et al. | Macrophage inflammatory protein-2: chromosomal regulation in rat small intestinal epithelial cells. | 1997 | Proc. Natl. Acad. Sci. U.S.A. | pmid:9294201 |
| Xu D et al. | Switch from Myc/Max to Mad1/Max binding and decrease in histone acetylation at the telomerase reverse transcriptase promoter during differentiation of HL60 cells. | 2001 | Proc. Natl. Acad. Sci. U.S.A. | pmid:11274400 |
| Boucher J et al. | Insulin and insulin-like growth factor 1 receptors are required for normal expression of imprinted genes. | 2014 | Proc. Natl. Acad. Sci. U.S.A. | pmid:25246545 |
| Chen WY et al. | Reactivation of silenced, virally transduced genes by inhibitors of histone deacetylase. | 1997 | Proc. Natl. Acad. Sci. U.S.A. | pmid:9159154 |
| Chen WY and Townes TM | Molecular mechanism for silencing virally transduced genes involves histone deacetylation and chromatin condensation. | 2000 | Proc. Natl. Acad. Sci. U.S.A. | pmid:10618426 |
| Zhang L et al. | Small molecule regulators of autophagy identified by an image-based high-throughput screen. | 2007 | Proc. Natl. Acad. Sci. U.S.A. | pmid:18024584 |
| Minucci S et al. | A histone deacetylase inhibitor potentiates retinoid receptor action in embryonal carcinoma cells. | 1997 | Proc. Natl. Acad. Sci. U.S.A. | pmid:9326603 |
| Duffy S et al. | Overexpression screens identify conserved dosage chromosome instability genes in yeast and human cancer. | 2016 | Proc. Natl. Acad. Sci. U.S.A. | pmid:27551064 |
| Matoba S et al. | RNAi-mediated knockdown of Xist can rescue the impaired postimplantation development of cloned mouse embryos. | 2011 | Proc. Natl. Acad. Sci. U.S.A. | pmid:22065773 |
| Crump NT et al. | Dynamic acetylation of all lysine-4 trimethylated histone H3 is evolutionarily conserved and mediated by p300/CBP. | 2011 | Proc. Natl. Acad. Sci. U.S.A. | pmid:21518915 |
| Plouffe D et al. | In silico activity profiling reveals the mechanism of action of antimalarials discovered in a high-throughput screen. | 2008 | Proc. Natl. Acad. Sci. U.S.A. | pmid:18579783 |
| Yeo M et al. | Bisphenol A delays the perinatal chloride shift in cortical neurons by epigenetic effects on the Kcc2 promoter. | 2013 | Proc. Natl. Acad. Sci. U.S.A. | pmid:23440186 |
| Saito A et al. | A synthetic inhibitor of histone deacetylase, MS-27-275, with marked in vivo antitumor activity against human tumors. | 1999 | Proc. Natl. Acad. Sci. U.S.A. | pmid:10200307 |
| Tavera-Mendoza LE et al. | Incorporation of histone deacetylase inhibition into the structure of a nuclear receptor agonist. | 2008 | Proc. Natl. Acad. Sci. U.S.A. | pmid:18550844 |
| Carmen AA et al. | Yeast HOS3 forms a novel trichostatin A-insensitive homodimer with intrinsic histone deacetylase activity. | 1999 | Proc. Natl. Acad. Sci. U.S.A. | pmid:10535926 |
| Ji Y et al. | A multistep mechanism for the activation of rearrangement in the immune system. | 2003 | Proc. Natl. Acad. Sci. U.S.A. | pmid:12802019 |
| Kim MY et al. | A repressor complex, AP4 transcription factor and geminin, negatively regulates expression of target genes in nonneuronal cells. | 2006 | Proc. Natl. Acad. Sci. U.S.A. | pmid:16924111 |
| Monte M et al. | MAGE-A tumor antigens target p53 transactivation function through histone deacetylase recruitment and confer resistance to chemotherapeutic agents. | 2006 | Proc. Natl. Acad. Sci. U.S.A. | pmid:16847267 |
| Furumai R et al. | Potent histone deacetylase inhibitors built from trichostatin A and cyclic tetrapeptide antibiotics including trapoxin. | 2001 | Proc. Natl. Acad. Sci. U.S.A. | pmid:11134513 |
| Subramanian C et al. | Ku70 acetylation mediates neuroblastoma cell death induced by histone deacetylase inhibitors. | 2005 | Proc. Natl. Acad. Sci. U.S.A. | pmid:15778293 |
| Li B et al. | FOXP3 interactions with histone acetyltransferase and class II histone deacetylases are required for repression. | 2007 | Proc. Natl. Acad. Sci. U.S.A. | pmid:17360565 |
| Zhou G et al. | HSV carrying WT REST establishes latency but reactivates only if the synthesis of REST is suppressed. | 2013 | Proc. Natl. Acad. Sci. U.S.A. | pmid:23341636 |