| MeSH term | MeSH ID | Detail |
|---|---|---|
| Hypertension | D006973 | 115 associated lipids |
| Hypertension, Pulmonary | D006976 | 32 associated lipids |
| Hypesthesia | D006987 | 1 associated lipids |
| Immunologic Deficiency Syndromes | D007153 | 8 associated lipids |
| Inflammation | D007249 | 119 associated lipids |
| Chromosome Inversion | D007446 | 1 associated lipids |
| Keloid | D007627 | 12 associated lipids |
| Leishmaniasis | D007896 | 19 associated lipids |
| Leukemia | D007938 | 74 associated lipids |
| Leukemia, Myeloid | D007951 | 52 associated lipids |
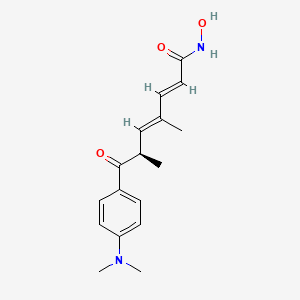
trichostatin A
Trichostatin is a lipid of Polyketides (PK) class. Trichostatin is associated with abnormalities such as Dentatorubral-Pallidoluysian Atrophy, PARAGANGLIOMAS 3, abnormal fragmented structure, Disintegration (morphologic abnormality) and Hyperostosis, Diffuse Idiopathic Skeletal. The involved functions are known as Acetylation, Cell Differentiation process, histone modification, Gene Silencing and Transcriptional Activation. Trichostatin often locates in CD41a, Hematopoietic System, Chromatin Structure, Blood and Endothelium. The associated genes with Trichostatin are SPI1 gene, CELL Gene, Chromatin, CXCR4 gene and DNMT1 gene. The related lipids are Butyrates, Promega, butyrate, Lipopolysaccharides and Steroids. The related experimental models are Knock-out, Mouse Model, Xenograft Model and Cancer Model.
Cross Reference
Introduction
To understand associated biological information of trichostatin A, we collected biological information of abnormalities, associated pathways, cellular/molecular locations, biological functions, related genes/proteins, lipids and common seen animal/experimental models with organized paragraphs from literatures.
What diseases are associated with trichostatin A?
trichostatin A is suspected in Infection, Morphologically altered structure, Ureteral obstruction, Photosensitization, Atherosclerosis, Hypertrophic Cardiomyopathy and other diseases in descending order of the highest number of associated sentences.
Related references are mostly published in these journals:
| Disease | Cross reference | Weighted score | Related literature |
|---|
Loading... please refresh the page if content is not showing up.
Possible diseases from mapped MeSH terms on references
We collected disease MeSH terms mapped to the references associated with trichostatin A
PubChem Associated disorders and diseases
What pathways are associated with trichostatin A
Lipid pathways are not clear in current pathway databases. We organized associated pathways with trichostatin A through full-text articles, including metabolic pathways or pathways of biological mechanisms.
Related references are published most in these journals:
| Pathway name | Related literatures |
|---|
Loading... please refresh the page if content is not showing up.
PubChem Biomolecular Interactions and Pathways
Link to PubChem Biomolecular Interactions and PathwaysWhat cellular locations are associated with trichostatin A?
Visualization in cellular structure
Associated locations are in red color. Not associated locations are in black.
Related references are published most in these journals:
| Location | Cross reference | Weighted score | Related literatures |
|---|
Loading... please refresh the page if content is not showing up.
What functions are associated with trichostatin A?
Related references are published most in these journals:
| Function | Cross reference | Weighted score | Related literatures |
|---|
What lipids are associated with trichostatin A?
Related references are published most in these journals:
| Lipid concept | Cross reference | Weighted score | Related literatures |
|---|
Loading... please refresh the page if content is not showing up.
What genes are associated with trichostatin A?
Related references are published most in these journals:
| Gene | Cross reference | Weighted score | Related literatures |
|---|
What common seen animal models are associated with trichostatin A?
Mouse Model
Mouse Model are used in the study 'Regulation of minichromosome maintenance gene family by microRNA-1296 and genistein in prostate cancer.' (Majid S et al., 2010), Mouse Model are used in the study 'Reversal of hypermethylation and reactivation of p16INK4a, RARbeta, and MGMT genes by genistein and other isoflavones from soy.' (Fang MZ et al., 2005) and Mouse Model are used in the study 'Histone deacetylase 3 mediates allergic skin inflammation by regulating expression of MCP1 protein.' (Kim Y et al., 2012).
Xenograft Model
Xenograft Model are used in the study 'Histone deacetylase inhibitors induce growth arrest and differentiation in uveal melanoma.' (Landreville S et al., 2012), Xenograft Model are used in the study 'Extended treatment with physiologic concentrations of dietary phytochemicals results in altered gene expression, reduced growth, and apoptosis of cancer cells.' (Moiseeva EP et al., 2007) and Xenograft Model are used in the study 'Retinoic acid and the histone deacetylase inhibitor trichostatin a inhibit the proliferation of human renal cell carcinoma in a xenograft tumor model.' (Touma SE et al., 2005).
Cancer Model
Cancer Model are used in the study 'Plasma pharmacokinetics and metabolism of the histone deacetylase inhibitor trichostatin a after intraperitoneal administration to mice.' (Sanderson L et al., 2004).
Related references are published most in these journals:
| Model | Cross reference | Weighted score | Related literatures |
|---|
Loading... please refresh the page if content is not showing up.
NCBI Entrez Crosslinks
All references with trichostatin A
Download all related citations| Authors | Title | Published | Journal | PubMed Link |
|---|---|---|---|---|
| Ji Z et al. | Integrating genomics and proteomics data to predict drug effects using binary linear programming. | 2014 | PLoS ONE | pmid:25036040 |
| Nishida N et al. | Identification of epigenetically inactivated genes in human hepatocellular carcinoma by integrative analyses of methylation profiling and pharmacological unmasking. | 2014 | Dig Dis | pmid:25376292 |
| Xie M et al. | Effects of 5-aza-2'-deoxycytidine and trichostatin A on high glucose- and interleukin-1β-induced secretory mediators from human retinal endothelial cells and retinal pigment epithelial cells. | 2014 | Mol. Vis. | pmid:25352747 |
| Quan H et al. | Hepatitis C virus core protein epigenetically silences SFRP1 and enhances HCC aggressiveness by inducing epithelial-mesenchymal transition. | 2014 | Oncogene | pmid:23770846 |
| Ishihara K | Eosinophil cell lines. | 2014 | Methods Mol. Biol. | pmid:24986606 |
| Liu H et al. | The Smn-independent beneficial effects of trichostatin A on an intermediate mouse model of spinal muscular atrophy. | 2014 | PLoS ONE | pmid:24984019 |
| Lee J et al. | Therapeutic targeting of epigenetic components in amyotrophic lateral sclerosis (ALS). | 2014 | Curr. Med. Chem. | pmid:25005187 |
| He G et al. | Inhibition of autophagy induced by TSA sensitizes colon cancer cell to radiation. | 2014 | Tumour Biol. | pmid:24122231 |
| Drela K et al. | Low oxygen atmosphere facilitates proliferation and maintains undifferentiated state of umbilical cord mesenchymal stem cells in an hypoxia inducible factor-dependent manner. | 2014 | Cytotherapy | pmid:24726658 |
| Zhu XJ et al. | Novel tumor-suppressor gene epidermal growth factor-containing fibulin-like extracellular matrix protein 1 is epigenetically silenced and associated with invasion and metastasis in human gastric cancer. | 2014 | Mol Med Rep | pmid:24718752 |
| Song BS et al. | Valproic acid enhances early development of bovine somatic cell nuclear transfer embryos by alleviating endoplasmic reticulum stress. | 2014 | Reprod. Fertil. Dev. | pmid:23506644 |
| Bitman M et al. | Valproate activates ERK signaling pathway in primary human hepatocytes. | 2014 | Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub | pmid:23073524 |
| Wei G et al. | [Functional analysis of Oct4 promoter in Xuhuai goat]. | 2014 | Yi Chuan | pmid:25143277 |
| Zou JY and Crews FT | Release of neuronal HMGB1 by ethanol through decreased HDAC activity activates brain neuroimmune signaling. | 2014 | PLoS ONE | pmid:24551070 |
| Guilluy C et al. | Isolated nuclei adapt to force and reveal a mechanotransduction pathway in the nucleus. | 2014 | Nat. Cell Biol. | pmid:24609268 |
| Cherry KE et al. | Identification of Tf1 integration events in S. pombe under nonselective conditions. | 2014 | Gene | pmid:24680781 |
| Yang J et al. | PTEN methylation involved in benzene-induced hematotoxicity. | 2014 | Exp. Mol. Pathol. | pmid:24680972 |
| Serrat N et al. | The response of secondary genes to lipopolysaccharides in macrophages depends on histone deacetylase and phosphorylation of C/EBPβ. | 2014 | J. Immunol. | pmid:24307736 |
| Hou SY et al. | Expressions of MAGE-A9 and MAGE-A11 in breast cancer and their expression mechanism. | 2014 | Arch. Med. Res. | pmid:24316396 |
| Reeves ME et al. | Evidence that RASSF1C stimulation of lung cancer cell proliferation depends on IGFBP-5 and PIWIL1 expression levels. | 2014 | PLoS ONE | pmid:25007054 |